ホーム > 砂糖 > 調査報告 > その他 > アセトン・ブタノール発酵の展望について〜糖質の多目的利用〜
アセトン・ブタノール発酵の展望について〜糖質の多目的利用〜
最終更新日:2010年3月6日
はじめに
バイオ燃料には広範な物性をもった植物由来の未利用資源が含まれますが、最もその利用が期待されているのは、バイオディーゼル(植物油およびそのエステル化物)、バイオエタノール、およびバイオブタノールです。バイオディーゼルやバイオエタノールは既に一部燃料としての使用が実現しています。素材そのものを使うバイオディーゼルに比べ、バイオエタノールとバイオブタノールは微生物による発酵工程を経て製造されます。食品として古代より作られたエタノール製造技術は既に成熟しており、砂糖や転化糖から「酵母」により生産されています。一方、バイオブタノールは、「アセトン・ブタノール菌」という細菌を用いて作られますが、エタノールに比べ水分を含み難く保存や流通に有利であること、単位容量あたりのエネルギー価が高くガソリン代替燃料として現在のエンジンにそのまま利用できることなど、その燃料特性が優れており、その効率的な生産法の開発が強く求められています。
バイオブタノールの歴史的背景
バイオ燃料が注目を浴びるようになった背景には、石油化学工業の負の面としての、化石燃料消費による地球環境汚染、特に地球温暖化の問題があります。また、化石燃料の枯渇問題と価格不安があります。特に近年、炭酸ガス放出による地球温暖化が緊要に解決すべき課題となっていますが、その主要な原因に自動車の排気ガスを挙げることができます。1900年代初頭、各種エンジンが開発されましたが、ディーゼルエンジンはピーナツオイルを、4サイクルエンジンはエタノールを燃料として使用できることが当時から知られていました。現在これらの再生利用可能資源を利用した燃料開発が改めて注目を浴びているのは面白いことです。あたかも自動車エンジンの開発者たちの所期の燃料に、現在に至って回帰しているかのようです。なお、この当時、ブタノールは燃料として使用されていません。というのも、当時はブタノールの大量生産法が開発されたばかりだったからです。
現在、ブタノールはエチレンから石油化学工業において生産されていますが、1900年代初頭、ブタノールはアセトン・ブタノール発酵によって生産が開始されました。このアセトン・ブタノール発酵は、合成ゴム製造原料としてのブタノール製造を目的として開発され、さらに第一次世界大戦中に、無煙火薬製造に不可欠のアセトンの製造法として大量生産法が確立しました。当時用いられた原料は、てん菜およびとうもろこしです。このように、アセトン・ブタノール発酵は化成品を発酵法で造る初の近代発酵工業として出発しました。当初、アセトンが目的産物であったため、ブタノールは副産物として利用法がありませんでした。しかし、その後、塗料や前述の合成ゴム原料として、ブタノールの有用性が認められるようになると、ブタノールが主要な目的産物となり、種々のアセトン・ブタノール菌が土壌から検索されるようになりました(1)。石油に乏しい日本ではバイオブタノールの航空燃料化も試みられたようですが、本格的利用とはならなかったようです。本発酵法は、第二次世界大戦後も廃糖みつを原料として、引き続き主要なブタノール生産法として存続しましたが、石油化学工業が隆盛するに従って競争力を失い、日本を含む主要工業国では1960年代までに消滅してしまいました。
アセトン・ブタノール発酵法を確立した人物としてワイツマンがよく知られていますが、彼はユダヤ人で、イギリス政府からアセトン・ブタノール発酵法の確立による功績をたたえられた際、個人としての名誉より、パレスチナにおけるユダヤ人国家の樹立を希望したそうです。第二次大戦後イスラエルが建国されると、彼は初代大統領となっています。皮肉なことに、石油へのエネルギー転換が起こって久しい70年代にイスラエルと周辺諸国とのあつれきによって勃発した中東戦争などにより石油価格が暴騰すると、いったん石油化学工業に席を譲ったアセトン・ブタノール発酵によるブタノール生産が改めて見直されるようになりました。
現在、アセトン・ブタノール発酵を行っている国として、イギリスと中国があります。イギリスでは、アメリカの化成品生産会社の雄として知られるDuPontと石油メジャーのBritish Petroleumのジョイントベンチャーが、ビートを原料とした試験的生産をおこなっています。中国では2004年までで終息したといわれていましたが、とうもろこしなどでん粉原料をもととした発酵製造が再開されています。
アセトン・ブタノール菌の性質
80年代になって再開されたアセトン・ブタノール発酵の研究は、先のアセトン・ブタノール発酵工業確立の時期の土壌からの新菌種の分離を主体としていた研究とは、様相が一変しました。その主要な研究課題は、アセトン・ブタノール発酵の効率化を目指すと共に、当時大腸菌を中心に開発が進んでいた遺伝子操作技術を用いた代謝の解析へと進んでいきました。
アセトン・ブタノール発酵菌は、少なくとも4種が存在することが現在明らかになっています(2)。アセトン・ブタノール発酵工業化初期に分離された菌株はClostridium acetobutylicumと呼ばれる細菌で、それ以後、世界各地で分離されてきた種々の菌株もC.acetobutylicumと類似した生理的挙動を示すので、これに類するものであろうと考えられていました。しかし、近年の遺伝子解析により多様な菌種が存在することがわかり、育種に必要な多様な遺伝子資源を提供できることが明らかになりました。他の3種はC.beijerinckii、C.saccharobutylicum、およびC.saccharoperbutylacetonicumで、特にC.saccharoperbutylacetonicumは本学の教授であった本江元吉により福岡市近郊で分離された菌株で、実際の工業生産に用いられたものです。
いずれの菌株も、エタノール生産に用いられる酵母と異なり、アミラーゼを生成することができますので、でん粉を直接発酵基質とした発酵生産が可能です。また、しょ糖、セロビオースなど2糖類、グルコースなど代表的な六単糖、さらに、いわゆる木質糖であるキシロース、アラビノースなどの五単糖やガラクトースも利用してアセトンやブタノールを生成することが出来ます。アセトン・ブタノール菌の有用性は、でん粉やグルコースなど動物飼料に用いられる糖類以外に広範な糖類を利用できるところにもあるのです。
面白いことに、これらの菌株は完全に酸素のないところでしか生育することが出来ません。酸素のあるところに菌株細胞をさらすと、アッという間に死んでしまいます。酸素に著しく弱いことから、これらの菌株による発酵生産が困難な工程であるかのように思えますが、実際に発酵生産を行う場合は、1ミリリットル中に数千万〜1億細胞程度という高い菌濃度の細胞を接種し、細胞自身が出す炭酸ガスや水素ガスにより、自らの成育環境を容易に酸素のない嫌気的環境に変えることができるようにします。そのため、むしろ酸素が必要な好気的細菌より簡便な培養装置で培養することが可能です。
また、酸素のない環境だからこそ、これらの菌はアセトンやブタノールを作るのです。というのも、好気性細菌や人間のように糖質のエネルギーを搾りきってしまうような代謝(呼吸)を行う場合では、糖質を完全に分解して炭酸ガスと水にしてしまい、後には何も残りません。アセトン・ブタノール菌は糖質から少量のエネルギーしか作ることが出来ないので、大量の糖質を取り込んで、大量の「廃棄物」を細胞外に放り出しています。その「廃棄物」がアセトンであり、ブタノールなのです。その他、炭酸ガスと水素ガスを生成しています。ブタノールが燃料として高い燃焼価を示すのは、原料の糖質からほとんどエネルギーを消費することなく作られているからです。
アセトン・ブタノール菌の研究課題
培養を始めると、アセトン・ブタノール菌は培養初期12時間程度の対数増殖期に盛んに細胞分裂して増殖し、その後緩やかな成長期(定常期)に入ります。本菌の特徴として、この生育期に依存した異なる生成物の生産という「生成物転換」があります。対数増殖期には酢酸と酪酸からなる脂肪酸を生成し、定常期になってアセトン、ブタノールおよび少量のエタノールを生成するようになるのです。この生成物転換がどうして、どのような機構で起こるのかが研究の興味のひとつです(図1)。
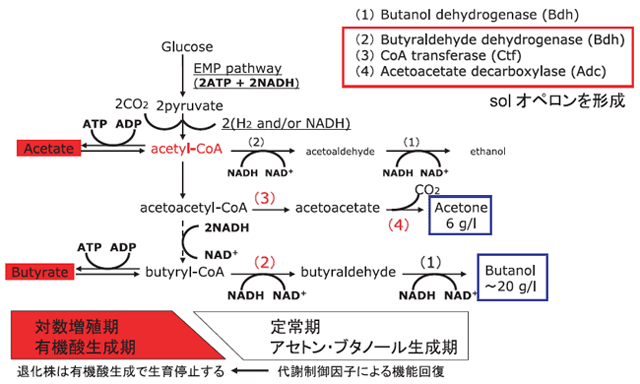 |
図1
アセトン・ブタノール菌の代謝経路の概略
|
アセトン・ブタノール菌を連続して培養し続けると、アセトン・ブタノール生産に重要な生成物転換が起こらない「退化」した菌株が生じて、アセトン・ブタノール発酵に大きな損害を与えることが知られています。物質生産を安定して行うためには、微生物の保存管理が重要な項目であることを示す事例です。「生成物転換」には種々の原因が考えられていますが、C.saccharoperbutylacetonicumでは、アセトン・ブタノール菌自身が、細胞外に「代謝転換因子」を蓄積し、さらに自ら代謝転換因子の蓄積量を検知して酸生成からアセトン・ブタノール生成へ代謝機能を転換していると考えられます(3)。
また、この代謝の大幅な転換を引き起こす本体として、solオペロンと呼ばれる一群の遺伝子があることがC.acetobutylicumで最初に見出されました。類似の遺伝子群が他のアセトン・ブタノール菌でも見つけられましたが、C.acetobutylicumとは少し異なっていることがわかりました。さらに、そのsolオペロンがC.acetobutylicumでは染色体外のプラスミド上に載っているのに、他の菌株では染色体上に載っていました。C.acetobutylicumでは、「退化」した菌株は、そのプラスミドを消失していることが明らかになっています(4)。C.acetobutylicumの「退化」の形式は染色体上にsolオペロンを有する他の3株とは異なるようです。
アセトン・ブタノール菌育種研究にとっての最大の課題は、菌株が生成するブタノール自身の毒性をどのようにして回避するかという点にあります。ほとんどのアセトン・ブタノール菌が1リットル当たり約15グラムのブタノールを生成すると発酵を停止してしまいます。現在までのチャンピオンデータでも同20グラムです(5)。この発酵上限は細胞膜に溶け込むブタノール濃度の上昇により細胞膜が破壊されるためと考えられており、ブタノール耐性を付与する遺伝子または物質の探索が続けられています。より高い濃度のブタノールに対する耐性が獲得されることは、発酵終了後のブタノール回収に大きな影響を与え、高濃度ブタノール生成ができれば回収に必要なエネルギーを大幅に削減することができるからです。発酵終了後のブタノールの回収法としては、蒸留法、膜透過法などが考えられますが、さらに培養中にブタノールを抽出してブタノール濃度を低減するなどの方法を組み合わせて、より効率的な回収法を開発することにより現在のブタノール生成限界によるデメリットを低減することができるようになるでしょう(図2)。
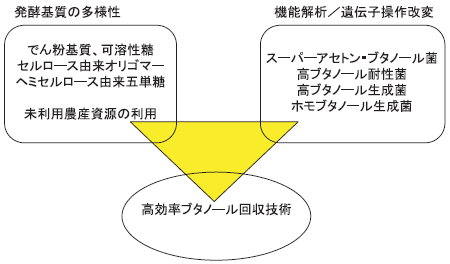 |
図2
アセトン・ブタノール菌の改良の展望
|
おわりに
アセトン・ブタノール菌がアセトン、ブタノール、エタノールなど複数の物質を生成するという性質は、生成物回収にとって困難な要因となります。ブタノールだけを発酵生産させるというホモブタノール生産は、自然界からの微生物分離を試みていた時代からの夢でもありますが、遺伝育種が可能になった現在、現実的な研究対象となっています。ホモブタノール生産では、代謝過程で生じた全ての還元力をブタノール生成のみに利用する菌株の育成が必要です。酵母で可能なホモエタノール生産と同じように、Clostridium属細菌でホモブタノール生産を実現することは必ずしも不可能ではないと思います。
さらに、最近になって、これらの遺伝子を大腸菌などの菌で発現させてブタノールを作らせようという試みがあります(6)。まだ試みの段階で、アセトン・ブタノール菌に比肩できる発酵能を示せる状態ではありませんが、ブタノールが持つ毒性に対抗できる性質を組み込みや、ホモブタノール生産への応用により、アセトン・ブタノール菌をしのぐ性質を持つ菌株の育種が期待されます。
文献
(1) Durre, P., Biobutanol: An attractive biofuel. Biotechnol. J. 2007, 2, 1525―1534.
(2) Keis, S., et. al., Emended descriptions of Clostridium acetobutylicum and
Clostridium beijerinckii, and descriptions of Clostridium saccharoperbutylacetonicum
sp. Nov. and Clostridium saccharobutylicum sp. Nov. Int. J. Syst. Evol. Microbiol.
2001, 51, 2095―2103.
(3) Kousaka, T., et. al., Characterization of the sol operon in butanol―hyper―producing
Clostridium saccharoperbutylacetonicum strain N1―4 and its degeneration mechanism.
Biosci. Bioctechnol. Biochem. 2007, 71, 58―68.
(4) Nolling, J. et. al., Genome sequence and comparative analysis of the solvent―producing
bacterium Clostridium acetobutylicum. J. Bacteriol. 2001, 183, 4823―4838.
(5) Qureshi, N. and Blaschek, H.P., Butanol production using Clostridium beijerinckii
BA101 hyper―butanol producing mutant strain and recovery by perevaporation.
Appl. Biochem. & Biotechnol., 1999, 84―86, 225―235.
(6) Atsumi, S. et. al., Metabolic engineering of Escherichia coli for 1―butanol
production. 2007, 10, 305―311.
| ページのトップへ |
このページに掲載されている情報の発信元
農畜産業振興機構 調査情報部 (担当:企画情報グループ)
Tel:03-3583-8713
農畜産業振興機構 調査情報部 (担当:企画情報グループ)
Tel:03-3583-8713